
What We Do
Background

Rita Levi-Montalcini
Source: http://livluna.com/rest-in-peace-rita-levi-montalcini/
In the vertebrate nervous system, billions of neurons must properly make trillions of connections with each other and peripheral targets in order to function. It is unlikely that neural connectivity is achieved through a “name-tag” mechanism whereby each neuron is programmed with a specific destination and synaptic partner. It is much more likely that these complex patterns of connectivity are governed by a broad series of rules, which we are only now beginning to understand. One of these rules can be inferred from the observation that during development, vertebrates over-produce neurons. As development progresses, a sculpting event occurs where roughly 50% of neurons die. Hamburger and Levi-Montalcini explained this cell loss with the neurotrophic factor hypothesis, which posits that neuronal survival during development is dependent on a limited supply of target-derived trophic factors. Although this may appear inefficient, a Darwinian-like competition between neurons for target-derived survival cues may represent an evolutionary solution for producing a functional nervous system with increased complexity and a limited number of instructive cues. My lab seeks to better understand this emergent property underlying neural development.
Summary of Significant Scientific Work
We recently found that developmental neuronal competition at the individual cell level involves a complex combination of events. We used mathematical modeling, biochemical, and genetic approaches to show that, in the PNS, the likelihood of neuronal survival is critically dependent on a sensitization process initiated by target-derived nerve growth factor (NGF) and strengthened by a series of positive-feedback loops (Science, 2008). We found that in addition to initiating expression of survival factors, the retrogradely transported NGF-TrkA signaling complex also increases expression of TrkA, thereby increasing the magnitude of NGF-TrkA pro-survival signaling. We also discovered that the duration of NGF-TrkA pro-survival signaling is variable, and regulated in an NGF-dependent manner via an unknown mechanism (Question1).
NGF also influences a complex protection/punishment mechanism affecting competition. We found that, in sympathetic neurons, NGF signaling induces neurons to increase secretion of unknown factors (Question 2) which, via p75 signaling, can kill or "punish" neighboring sympathetic neurons in a paracrine manner. High NGF-TrkA signaling protects neurons from this punishment cue while low NGF-TrkA signaling leaves cells vulnerable. In my lab we are working to not only understand the molecular basis for these competitive processes (Questions 1 and 2), we are also examining how this logic applies to other aspects of neural development, function, and disease (Questions 3-5). Finally we are working to develop new tools to interrogate nervous system function (Question 6). We are currently addressing the following questions:
1. How is the duration of long-distance trophic signaling regulated? Communication from axonal targets to neuronal cell bodies is critical for both the development and function of the nervous system. How this retrograde long-distance signal maintains high fidelity as it traffics back to the cell body remains unknown. Current Work: Using novel tools to track post-endocytic NGF-TrkA (Neuron, 2010) we found that the retrograde signaling endosome induces the expression and recruitment of a novel effector protein, Coronin-1, which protects the endosome from lysosomal degradation. The mechanism by which Coronin-1 protects the signaling endosome is by facilitating NGF-dependent calcium release and subsequent calcineurin activation. Loss of Coronin-1 impairs several NGF-dependent processes including calcium release and neuron survival. This novel feedback loop mediating NGF-TrkA endosome stability represents a critical, missing component of our model of developmental competition for survival, which is implicit within the neurotrophic factor hypothesis (Nat. Neuro, 2014). We also found that Coronin-1 acts as a molecular switch to change the way in which axons behave when the innervate final target organs like the eye or the heart (J. Neuro, 2015). We are also examining how various neurotrophic factors use coronin-1 to facilitate other competitive developmental events such as synapse formation. In collaborate on this project with Prof. Bettina Winkler we are seeking a better understanding of the role of coronin-1 in signaling endosome trafficking and function.
2. What is the relationship between protection and punishment signaling pathways? All of neural pattern formation requires signals that induces building or destruction. Implicit within this idea is an antagonistic relationship between pro-growth and pro-refinement pathways. These antagonistic pathways often emanate from receptor tyrosine kinases (RTKs) as pro-growth and tumor necrosis factor receptor (TNFR) family members as pro-refinement. The balance between these growth and refinement cues defines the structure of the nervous system, yet very little is known about cross-talk between these signals. We are now seeking to understand the crosstalk not only between RTKs and TNFR family members but also how TNFR family members work with each other to sculpt the nervous system
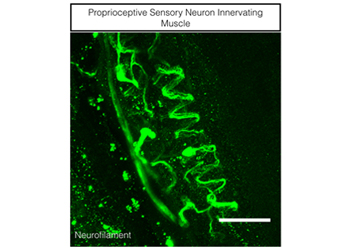
3. How does punishment signaling influence cutaneous sensory perception? We have begun to consider how developmental competition pathways are reused in adults. It is well established that overstimulation or understimulation of the TrkA RTK underlies several pain conditions in humans. If the aforementioned functional antagonism between RTK and TNFR pathways is recycled for non-developmental functions, it would stand to reason that gain or loss of function of TNFR family members would also display a pain phenotype. Indeed, we recently found a novel antagonistic interaction between TNFR1 and TrkA to govern sensitivity to touch (Neuron, 2014). This insight may allow us to better rationalize pharmacological interventions for chronic pain disorders which impact one quarter of the World’s population. We are now examining how these antagonistic interactions buffer other sensory modalities like proprioception.
4. Are competitive signaling pathways re-engaged to facilitate post-developmental homeostatic changes in metabolism? We have begun to consider how the sympathetic nervous system uses protection and punishment pathways to properly respond to changes in diet which in turn regulates energy storage/utilization to-and-from fat stores. We are broadly interested in understanding how the sympathetic nervous system regulates energy liberation from fat in response to particular diets. We are also interested in how the developmental antagonistic signaling pathways mentioned above may regulate this.
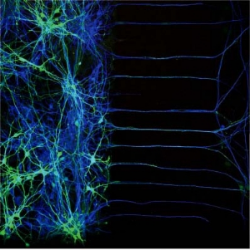
Taken in collaboration with Nikhil Sharma and David Ginty.
5. What common pathways underly developmental, injury-, and disease-mediated axon degeneration? The competitive signaling programs described above are widely appreciated to be involved in a type of developmental axon degeneration called pruning. Pruning shares several morphological similarities with both disease and injury induced degeneration. Because of these morphological similarities, we argue that they may share common molecular mechanisms despite having different etiologies. By understanding the commonalities between different types of degeneration, we hope to better rationalize treatments for neurodegenerative disease and nerve injury.
6. Technology Development: Magnetogenetics and Rapid Phenotyping via Mass Cytometry: We are also building tools, which will enable next generation experiments related to development, function and pathology of the nervous system. 1. Magnetogenetics: In collaboration with Ali Güler’s lab, we are building non-invasive magnetogenetic tools to better understand the signaling pathways underlying neural circuit assembly and function. The first of these tools, Magneto, is capable of remotely inducing the firing of neurons in awake and behaving animals (Nat. Neuro, 2016; 2019). 2. Mass Cytometry. In collaboration with Eli Zunder’s lab, we are developing a method to rapidly characterize the composition and signaling of cells in the nervous system. Using Mass cytometry we can characterize 50 markers at once in a single cell and millions of cells can be analyzed within hours. This emergent technology will illuminate new developmental differentiation pathways in the PNS (Nature Neuro, 2022) and CNS (Biorxv, 2022) and will allow for the rapid phenotyping of genetically modified mice.